В исследовании приняло участие 20 здоровых мужчин в возрасте 19–21 года. Условия проведения эксперимента были согласованы с комитетом по биоэтике ВЛГАФК, все испытуемые получили подробную информацию о проводимом исследовании и дали письменное согласие на участие в нем в соответствии с Хельсинкской декларацией.
Регистрация изометрических, концентрических и эксцентрических сокращений мышц голени исследуемых проводилась на мультисуставном комплексе Biodex Multi-Joint System Рго-3 (USA, 2006). Исследуемые выполняли плантарное и дорсальное сгибание стопы в голеностопном суставе в положении сидя. В первой части опытов испытуемые выполняли однократные сокращения в 50 % от максимального произвольного сокращения (МПС), во второй части – многократные сокращения в 80 % МПС (2 подхода по 6 повторений в каждом). Траектория движения в голеностопном суставе в обеих сериях составляла 45°, а продолжительность – 2 с. Изометрическое сокращение осуществлялось при трех суставных углах – 102°, 83° и 63°.
Регистрация электромиографической активности мышц (ЭМГ) осуществлялась по традиционной методике (Р.С. Персон, 1969; Р.М. Городничев, 2005) при помощи 8-канального электронейромиографа "Нейро-МВП-8" (ООО "Нейрософт", Россия, 2006) с использованием биполярных поверхностных (накожных) электродов. Определялись амплитуда и число турнов (поворотов) ЭМГ. ЭМГ регистрировалась с икроножной, камбаловидной и передней большеберцовой мышц. Полученные ЭМГ-записи обрабатывались в компьютерной программе "НейроМВП".
Транскраниальная магнитная стимуляция (ТМС) наносилась на головной мозг в области представительства исследуемых мышц с помощью магнитного стимулятора Magstim Rapid 2 (UK, 2007) с использованием угловой катушки диаметром 110 мм. Тестирующий магнитный стимул составлял расчетную величину: порог вызванного моторного ответа (ВМО) χ 1,2. Для возбуждения нейронов двигательной зоны головного мозга катушка располагалась в области проекции исследуемой мышцы в моторной коре левого полушария. За пороговое значение принималась сила магнитного стимула, при котором регистрировался ВМО с амплитудой не менее 0,1 мВ. Порог измеряли в % от выходной мощности магнитного стимулятора и в абсолютных значениях – теслах (Т). Оценивали амплитуду ВМО и корковый период молчания ЭМГ исследуемых мышц при ТМС. Амплитуда ВМО скелетных мышц определялась от пика до пика, длительность коркового периода молчания (КПМ) измерялась от момента окончания ВМО до конца абсолютного биоэлектрического молчания ЭМГ. По длительности периода молчания ЭМГ судили о выраженности тормозных процессов спинального и коркового уровня (R.M. Naatanen, 1987; M. Inghilleri et al., 1992; K. Nakazawa et al., 1997). Принималось: чем больше длительность периода молчания, тем больше выраженность (проявление) тормозных процессов.
В первой части опытов при однократном выполнении концентрического и эксцентрического сокращения амплитуда ВМО и КПМ исследуемых мышц определялись в начале, середине и при окончании сокращения. Регистрация ВМО осуществлялась также и в состоянии покоя. При выполнении изометрических сокращений эти параметры определялись в момент кратковременного удержания достигнутого статического усилия.
Во второй части опытов при многократном (2 подхода по 6 повторений в каждом) выполнении изометрических, концентрических и эксцентрических сокращений ВМО и КПМ исследуемых мышц регистрировались в первом, третьем и шестом повторении каждого подхода. ВМО и период молчания во всех случаях определялись в середине концентрического и эксцентрического типов сокращения и в момент достижения плато изометрического напряжения скелетных мышц. Заметим, что в протоколе 2-й части экспериментов выполняемые двигательные действия, количество подходов и повторений, а также периоды отдыха между повторными движениями моделировали реальную программу силовой тренировки.
Экспериментальные факты, полученные в 1-й части исследования, показали, что длительность периода молчания, а следовательно, и проявление корковых и сегментарных тормозных процессов, воздействующих на мышцы-агонисты, изменяются в зависимости от типа мышечного сокращения (табл. 6.1).
Таблица 6.1
Длительность периода молчания мышц-агонистов при различных типах мышечного сокращения, М ± m, n = 12
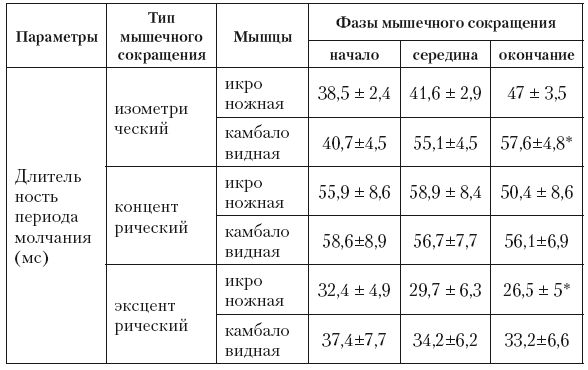
Примечания: * Р<0,05 – достоверность различий между значениями, зарегистрированными в начале и при окончании сокращения; при изометрическом сокращении в фазах – начало, середина и окончание – приведены величины периода молчания, полученные при суставном угле 102°, 83° и 63°.
Тормозные процессы при эксцентрическом сокращении были более слабыми в сравнении с их проявлением в ходе сокращений других типов. Они прогрессивно возрастали с уменьшением длины мышцы при изометрическом сокращении, понижались к моменту окончания эксцентрического сокращения и не менялись в разные фазы сокращения концентрического типа. Эти факты указывают на специфику процессов торможения активности а-мотонейронов, развивающихся на спинальном и корковом уровнях ЦНС, при мышечных сокращениях разного типа.
Амплитуда ВМО мышц-агонистов возрастала в процессе всех трех типов сокращений по сравнению с исходными значениями (рис. 6.3). Как видно из рисунка 6.3, наибольший прирост амплитуды ВМО по отношению к фоновым значениям отмечался в середине концентрического сокращения камбаловидной мышцы. Он превышал показатели, полученные при выполнении эксцентрической активности, на 495,6 %; изометрической – на 640,7 % (в обоих случаях Р<0,05).
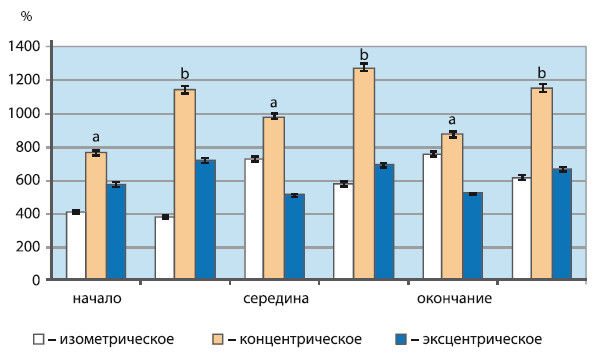
Рис. 6.3. Амплитуда ВМО скелетных мышц при различных типах мышечного сокращения по отношению к фоновым значениям (в %)
По оси абсцисс – фазы мышечного сокращения; a – икроножная мышца; b – камбаловидная мышца. При изометрическом сокращении в фазах – начало, середина и окончание – приведены значения амплитуды ВМО, полученные при суставном угле 102°, 83° и 63°
Известно, что первая треть КПМ обусловлена комбинацией корковых и спинальных механизмов, из которых большую значимость приобретают механизмы, обеспечивающие рефрактерность спинальных мотонейронов и возвратное торможение клеток Реншоу, а все остальное время исчезновения биоэлектрической активности обеспечивается корковым подавлением (K.R. Mills et al., 1987). Исходя из этого, можно предположить, что увеличение продолжительности КПМ скелетных мышц во время однократных сокращений концентрического и изометрического типа по сравнению с эксцентрическим связано с усилением процессов подавления активности мотонейронов, развивающихся на спинальном и корковом уровнях. Большее повышение амплитуды ВМО при концентрическом сокращении в сравнении с другими его типами свидетельствует о более значительном повышении возбудимости корковых нейронов при таком типе активности.
Сравнительный анализ результатов исследования выраженности корковых и спинальных тормозных процессов в ходе многократного выполнения (2 подхода по 6 повторений в каждом) мышечных сокращений различного типа выявил прогрессивное увеличение торможения от первого повторения к шестому в обоих подходах при изометрическом и эксцентрическом сокращениях мышц-агонистов. При выполнении повторных концентрических мышечных сокращений статистически значимых изменений периода молчания не происходило. Эффект увеличения тормозных процессов, воздействующих на икроножную мышцу и отражающихся в возрастании продолжительности периода молчания, представлен в таблице 6.2.
Таблица 6.2
Период молчания (мс) икроножной мышцы при ТМС в процессе многократных мышечных сокращений разного типа,
M± m, n=12