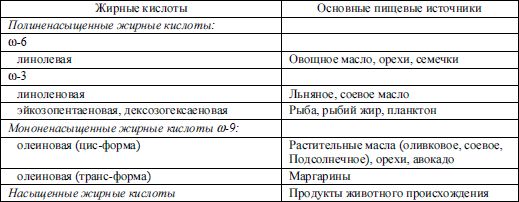
Особое значение в диетологии придается ненасыщенным жирным кислотам, являющимся эссенциальными факторами питания. Полиненасыщенные жирные кислоты, несущие в организме важнейшие функции (это предшественники ряда биологически активных веществ), должны поступать экзогенно. К эссенциальным жирным кислотам относятся линолевая и линоленовая. Линолевая кислота метаболизируется в организме в арахидоновую, а линоленовая в эйкозапентайеновую кислоты, которые могут поступать в организм с мясными и рыбными продуктами, но в незначительных количествах (табл. 3.5), и являются компонентами клеточных мембран, предшественниками гормоноподобных веществ. Линолевая и образуемая из нее арахидоновая кислота относятся к ω-6- жирным кислотам , линоленовая кислота и продукты ее метаболизма эйкозо-пентаеновая и дезоксогексаеновая – ω-3 -жирные кислоты. Дефицит эссенциальных жирных кислот в рационе вызывает, прежде всего, нарушение биосинтеза арахидоновой кислоты, которая входит в большом количестве в состав структурных фосфолипидов и простагландинов. Содержание линолевой и линоленовой кислот во многом определяет биологическую ценность пищевых продуктов. Недостаточность эссенциальных жирных кислот развивается в основном у больных, находящихся на полном парентеральном питании без применения жировых эмульсий.
При обработке растительных масел – создании маргаринов – происходит изомеризация ненасыщенных жирных кислот с созданием трансизомеров, которые утрачивают некоторые биологические функции своих предшественников.
Триглицериды со средней длиной углеродной цепи (СЦТ) имеют более высокую усвояемость, чем другие виды триглицеридов. Они гидролизируются в кишечнике без участия желчи, больше атакуются липазами. Кроме того, введение среднецепочечных триглицеридов оказывает гипохолестеринемический эффект, так как они не участвуют в мицеллообразовании, необходимом для всасывания холестерина. Недостатком применения препаратов, содержащих триглицериды со средней длиной углеродной цепи, является то, что они используются исключительно как энергетический (но не пластический) субстрат. Кроме того, окисление таких жирных кислот приводит к интенсивному накоплению кетоновых тел и может усугубить ацидоз.
Таблица 3.5 Основные пищевые источники различных жирных кислот
Стерины и фосфолипиды не относятся к эссенциальным факторам питания, но играют важнейшую роль в метаболизме.
Фосфолипиды являются незаменимыми компонентами организма. Основная их роль – обеспечение фундаментальной структуры мембраны, как барьера проницаемости. Биосинтез структурных фосфолипидов в печени направлен на обеспечение ими самой печени и других органов. Фосфолипиды оказывают липотропное действие, способствуя мицеллообразованию жиров в пищеварительном тракте, транспорту их из печени, а также и стабилизируя липопротеины.
Стерины в животных продуктах представлены холестерином, а в растительных – смесью фитостеринов.
Холестерин является структурным компонентом мембран и предшественником стероидов (гормонов, витамина D, желчных кислот). Пополнение запасов холестерина происходит за счет кишечной абсорбции и биосинтеза (1 г в сутки). Количество всасывающегося в кишечнике холестерина ограничено (0,3–0,5 г в сутки), и при излишнем содержании в пище он выводится с фекалиями. Абсорбция холестерина ингибируется его растительными структурными аналогами фитостеринами. Сами фитостерины тоже могут включаться в эндогенные липидные образования, но их участие минимально. При избыточном поступлении холестерина с пищей его синтез в печени, кишечнике и коже практически прекращается.
Поступающий из кишечника в составе хиломикронов холестерин в значительной степени задерживается в печени, где используется для построения мембран гепатоцитов и в синтезе желчных кислот. В составе желчи в результате реабсорбции в организм возвращается около 40 % жиров. Не подвергшиеся обратному всасыванию в кишечнике холестерин и желчные кислоты – это основной путь выведения холестерина из организма. В кровотоке липиды существуют в составе транспортных форм: хиломикронов, липопротеидов очень низкой плотности (ЛПОНП), липопротеидов низкой плотности (ЛПНП) и липопротеидов высокой плотности (ЛПВП). В энтероцитах образуются хиломикроны и ЛОПНП, в гепатоцитах – ЛПОНП и ЛПВП, в плазме крови – ЛПВП и ЛПНП (рис. 3.3).
Хиломикроны и ЛПОНП транспортируют преимущественно триглицериды, а ЛПНП и ЛПВП – холестерин. Холестерин-содержащие липопротеиды регулируют баланс холестерина в клетках: ЛПНП обеспечивают потребности, а ЛПВП предупреждают избыточное накопление.
Различают пять типов дислипопротеинемий. I тип связан с нарушением лизиса хиломикронов, Па тип – результат нарушения распада ЛПНП и снижения поступления холестерина в клетку, II тип характеризуется замедлением распада ЛПОНП, IV тип связан с усилением синтеза триглицеридов в печени в результате гиперинсулинизма, механизмы развития Пб и V типов точно не известны.
На состав триглицеридов и липопротеинов выраженное влияние оказывает состав пищи. Продукты животного происхождения, включающие преимущественно полиненасыщенные жирные кислоты и холестерин, имеют атерогенный эффект, влияют на содержание в крови ЛПВП и триглицеридов. Наоборот, ненасыщенные жирные кислоты (их источник растительные масла) и в особенности ω-3-жирные кислоты (содержащиеся в жире рыб) оказывают профилактическое действие (табл. 3.6).
Таблица 3.6 Влияние жирных кислот на липопротеидный спектр
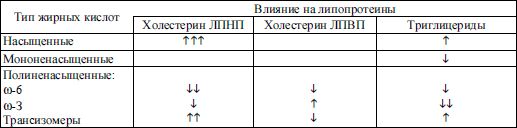
Примечание: ↑ – повышают, ↓ – снижают.

Рис. 3.3. Обмен холестерина (ХС) (по: Arias I. М. et al., 1982) (схема).
Как и при метаболизме углеводов, ведущую роль в липидном обмене играет печень. Исключительно в печени локализованы такие процессы, как биосинтез холестерина, желчных кислот и фосфолипидов. В обмене других липидов ей присущи модифицирующие и регуляторные функции. В отличие от богатых запасов гликогена, печень практически не содержит собственных резервов триглицеридов (менее 1 %), однако занимает ключевую позицию в процессах мобилизации, потребления и синтеза жиров в других тканях. Такая ее роль основана на том, что практически все потоки обмена жиров проходят через печень: липиды пищи в виде хиломикронов поступают в нее через общий кровоток по печеночной артерии; свободные жирные кислоты, мобилизованные из жировых депо, переносятся в виде комплексов с альбумином; соли желчных кислот, реабсорбируясь в кишечнике, вновь поступают по воротной вене.
Энергетический потенциал липидов обеспечивает более половины основной энергетической потребности большинства тканей, что особенно выражено в условиях голода. При голодании или снижении утилизации глюкозы, триглицериды жировой ткани гидролизируются в жирные кислоты, которые в таких органах, как сердце, мышцы и печень подвергаются интенсивному (β-окислению с образованием АТФ.
Продуктами неполной утилизации жиров печенью являются кетоновые тела. К ним относятся ацетоуксусная кислота, (β-оксибутират и ацетон. В норме кетоны образуются в незначительном количестве и полностью утилизируются как источник энергии нервной тканью, скелетными и висцеральными мышцами. В условиях ускоренного катаболизма жирных кислот и/или снижения утилизации углеводов синтез кетонов может превысить возможности их окисления внепеченочными органами и привести к развитию метаболического ацидоза. Ингибирующее влияние на кетоногенез оказывают углеводы рациона.
Головной мозг и нервная ткань практически не используют жиры как источник энергии, так как здесь не происходит (β-окисления. Однако эти ткани могут использовать кетоновые тела. В норме доля процессов окисления кетоновых тел незначительна по сравнению с катаболизмом глюкозы. Однако в условиях голодания кетоновые тела становятся важным альтернативным источником энергии.
Кетоны используются и мышцами , наряду с происходящей здесь утилизацией глюкозы и (β-окислением. При незначительной физической нагрузке мышцы окисляют в основном углеводы, увеличение интенсивности и длительности работы требует преобладания катаболизма жиров, (β-окисление в большинстве тканей стимулируется переносчиком липидов карнитином, но особенно весомое значение он имеет для мышечной ткани.
Свободнорадикальные формы кислорода вызывают процессы перекисного окисления, которому в первую очередь подвержены полиненасыщенные жирные кислоты. Это физиологический процесс, обеспечивающий регуляцию активности клеток. Однако при избыточном образовании свободных радикалов их окислительная активность приводит к нарушению структуры и гибели клетки. Для ограничения перекисного окисления существует система антиоксидантной защиты, которая ингибирует образование свободных радикалов и разлагает токсичные продукты их окисления. Функционирование этой системы во многом зависит от алиментарно поступающих антиоксидантов: токоферолов, селена, серосодержащих аминокислот, аскорбиновой кислоты, рутина.
Синтез жирных кислот (за исключением эссенциальных) может происходить из любых веществ, для которых конечным продуктом метаболизма является ацетил-КоА, но основным источником липогенеза являются углеводы. При излишнем количестве глюкозы в печени (после еды) и достаточных запасах гликогена глюкоза начинает разлагаться до предшественников жирных кислот. Если потребление углеводов превышает энергетические потребности организма, то их избыток в дальнейшем превращается в жиры.