T.M. Manini с коллегами (2007) определили относительную силу мышц, обеспечивающих разгибание в коленном суставе, чтобы оценить степень риска возможного ограничения двигательной активности у пожилых лиц (возраст 70–79 лет). Оказалось, что женщины и мужчины с относительной силой в диапазоне 1,01-1,13 Н х м/кг имели высокую степень риска, а обследуемые с величинами от 1,34 до 1,71 Н х м/кг – невысокий риск ограничения их двигательной активности. В другой работе исследовали взаимосвязь между уровнем развития силы разгибателей ноги и сохранением равновесия в группе молодых (средний возраст 21 год), среднего возраста (40 лет) и пожилых (71 год) мужчин (М. Izquierdo et al., 1999). Авторы выявили, что невысокая способность быстро развивать силу у пожилых людей связана с понижением нервно-мышечной регуляции устойчивости вертикальной позы. У пожилых людей в период наибольшего количества падений также исследовали силу нижних конечностей и величину максимального вращательного момента как факторов, ограничивающих равновесие (М. Pijnappels et al., 2008). Оказалось, что величина измеренной максимальной изометрической силы позволяет точно дифференцировать пожилых людей, с которыми случались или нет падения. В недавних исследованиях ассиметрия силы мышц нижних конечностей была определена как фактор риска падений у пожилых (средний возраст 74 года) женщин (D.A. Skelton et al., 2002). Аргументируется, что ассиметрия активности мышц нижних конечностей, вероятно, больше всего наносит ущерб регуляции центра массы тела над ограниченной поверхностью опоры (Е. Pоrtegijs et al., 2006).
Механизмы снижения силовых способностей у людей пожилого возраста. Точно установлено наличие связи между величиной массы скелетной мышцы и силой, которую она способна генерировать (D.T Kirkendall, W.E.Garrett, 1998; L.M. Maltais et al., 2015). Следовательно, уменьшение массы скелетных мышц при старении должно определять связанную с возрастом потерю силы. Действительно, проведенные исследования выявили снижение массы скелетных мышц на 30–50 % у женщин и мужчин в период от 40 до 80 лет (J. Lexell et al., 1988; Н.Н. Акша et al., 2001). При исследовании методом магнитного резонанса разнородных образцов мышц у 468 мужчин и женщин в возрасте 18–88 лет установлено понижение скелетной мышечной массы тела, начинавшееся после достижения 30-летнего возраста (I. Janssen et al., 2000). Однако значительное ее снижение происходило лишь в конце пятого десятилетия. Потеря мышечной массы всего тела не зависела от его длины и была значительнее у мужчин, чем у женщин в абсолютных и относительных величинах по отношению к массе тела. Кроме того, наблюдалось более значительное снижение массы скелетных мышц с возрастом в нижней части тела и у мужчин и у женщин. Авторы высказали предположение, что мышцы нижних конечностей требуют большей двигательной активности (ходьбы, подъема на ступеньки), поэтому тот факт, что потери мышечной массы преобладают в нижней части тела, вероятно, может быть объяснен главным образом связанным с возрастом понижением физической активности.
Понижение мышечной массы при старении зависит от снижения общего числа мышечных волокон, уменьшения их объема и комбинаций этих двух факторов (J.A. Faulkner et al., 2007). Определенное значение в снижении мышечной массы также имеют изменения внутреннего расположения мышечных волокон (мышечной архитектуры).
Снижение мышечной массы происходит в меньшей степени, чем мышечной силы (А. Дж. Мак-Комас, 2001). В некоторых мышцах уменьшение объема волокон может существенно превышать процентное снижение площади поперечного сечения мышцы вследствие замены сократительных элементов жировой и соединительной тканями. Такая замена мышечной ткани объясняет причину меньшей величины усилия, развиваемого пожилыми людьми на единицу площади поперечного сечения мышцы, по сравнению с молодыми (A.A. Vandervoort, AJ. McComas, 1986).
M.V. Narici с соавторами (2003) исследовали длину пучка икроножной мышцы, а также угол, под которым пучки вставляются в ее жилистую оболочку в группе молодых (возраст 27–42 года) и пожилых (возраст 70–81 год) физически активных мужчин. Выявлено, что пучки икроножной мышцы пожилых людей были короче на 10 % в сравнении с молодыми исследуемыми. Кроме того, величина наклона пучков к жилистой оболочке была на 13 % меньше у пожилых лиц. Эти результаты определяются связанной с возрастом потерей саркомеров в системе их последовательных соединений. По данным M.V. Narici с коллегами (2003), такое снижение числа саркомеров приводит к ослаблению функции мышцы у пожилых людей, влияя на зависимость следующих взаимоотношений соответствующей мышцы: длина – сила; сила – скорость и мощность – скорость. Результаты старения проявляются не только в потере саркомеров в последовательных соединениях, но также в уменьшении саркомеров в параллели.
Понижение массы мышц до 50-летнего возраста зависит главным образом от уменьшения поперечного сечения отдельных волокон вследствие малоподвижного образа жизни (J. Lexell, С.С. Taylor, 1991). На более поздних этапах жизни дополнительная потеря мышечной массы определяется уменьшением числа мышечных волокон. J. Lexell с соавторами (1988) наблюдали уменьшение количества мышечных волокон в наружной широкой мышце бедра на 50 % у пожилых людей (возраст 50–80 лет). Снижение числа мышечных волокон в пожилом возрасте сопровождается инфильтрацией соединительной и жировой ткани (E. Edstrom, B. Ulfhake, 2005), а также изнашиванием сателитных клеток. V. Renault с коллегами (2002) отметили понижение пропорции сателитных клеток на 1,44-1,77 % у пожилых людей (возраст 74 года) по сравнению с молодыми (средний возраст 23 года). Понижение числа мышечных волокон и сателитных клеток у пожилых лиц происходит вследствие повышенной чувствительности их клеточных структур к процессу гибели ядер без потери самих клеток (S.E. Alway, Р.М. Siu, 2008). Уменьшение числа сателитных клеток оказывает двойной негативный эффект на мышечные волокна, приводящий к их старению: ослабление регенеративного потенциала скелетной мышцы и уменьшение способности стареющей мышцы отвечать гипертрофией на адекватные тренировочные нагрузки.
Вопрос о типе мышечных волокон, ответственных за нервномышечное старение, является довольно сложным. Исследования показывают, что волокна типа II более подвержены процессам старения, чем волокна типа I (J. Lexell et al., 1988; A.R. Coggan et al, 1992) (рис. 8.1).
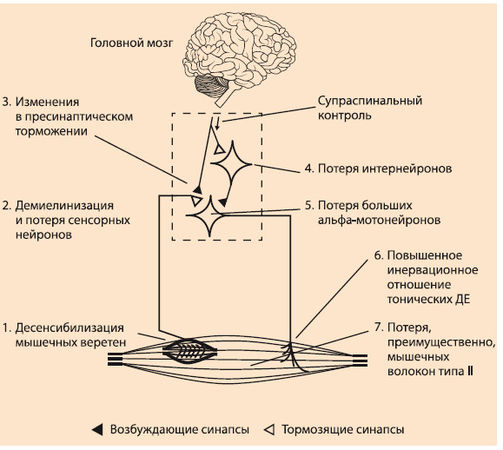
Рис. 8.1. Функциональные изменения моторной системы человека при старении
J. Lexell с соавторами (1988) наблюдали понижение на 26 % размера волокон типа II в наружной широкой мышце бедра от 20 лет к 80 годам. Изучение методом биопсии икроножной мышцы у молодых (средний возраст 24 года) и пожилых (средний возраст 64 года) мужчин и женщин показало снижение на 13–31 % поперечной площади волокон типа 11а и 11в (A.R. Coggan et al., 1992).
Могут быть и другие механизмы, ответственные за понижение числа и размера скелетных мышечных волокон в пожилом возрасте. Понижение физической активности приводит к уменьшению размера мышцы, тогда как процесс гибели ядер в мышечных волокнах и разрушение нейронального входа в мышцу являются причиной понижения мышечных волокон типа II (T. J. Doherty, 2003; S.E. Alway, P.M. Siu, 2008). Уменьшение числа быстрых мышечных волокон является причиной последовательного устранения больших альфа-мотонейронов (S. Тегао et al., 1996). Снижение количества быстрых мотонейронов приводит к денервированию волокон типа II. Некоторые волокна становятся реинервированными аксональными ответвлениями небольших альфа-мотонейронов (M.R. Roos et al., 1997). В результате этого просходит увеличение иннервационного отношения в оставшихся медленных двигательных единицах (M.R. Roos et al., 1997). Волокна, которые не становятся ренеиннервируемыми, остаются денер-вированными и со временем погибают.
Исследования последних лет показали, что не потеря мотонейронов ответственна за снижение мышечной массы, а нарушение способности волокна к регенерации и реиннервации (Е. Ed-strom et al, 2007). Это аргументируется тем, что связанная с возрастом потеря мотонейронов относительно мала (10–15 %) и что такое понижение не может в полной мере объяснить уменьшение значительного количества мышечных волокон (В. Ulfhake et al., 2000). Кроме того, уменьшение количества и размера мышечных волокон скелетных мышц является также следствием регенерации самих волокон (Е. Edstrom, В. Ulfhake, 2005). Авторы предполагают, что хотя мотонейроны денервируют часть мышечных волокон, тем не менее через некоторое время интактные отростки этих нейронов пытаются реиннервировать разъединенные мышечные волокна. Так как пластичность моторных аксонов уменьшается при старении (В.М. Carlson, J.A. Faulkner 1998), то невозможность реиннервирования свободных волокон, в конечном итоге, может приводить к снижению общего количества мышечных волокон (Е. Edstrom et al., 2007).