Регуляция ЦТД.
Осуществляется с помощью дыхательного контроля.
Дыхательный контроль – это регуляция скорости переноса электронов по дыхательной цепи отношением АТФ/АДФ. Чем меньше это отношение, тем интенсивнее идет дыхание и активнее синтезируется АТФ. Если АТФ не используется, и его концентрация в клетке возрастает, то прекращается поток электронов к кислороду. Накопление АДФ увеличивает окисление субстратов и поглощение кислорода. Механизм дыхательного контроля характеризуется высокой точностью и имеет важное значение, так как в результате его действия скорость синтеза АТФ соответствует потребностям клетки в энергии. Запасов АТФ в клетке не существует. Относительные концентрации АТФ/АДФ в тканях изменяются в узких пределах, в то время как потребление энергии клеткой может изменяться в десятки раз.
Американский биохимик Д. Чанс предложил рассматривать 5 состояний митохондрий, при которых скорость их дыхания ограничивается определенными факторами:
1. Недостаток SH2 и АДФ – скорость дыхания очень низкая.
2. Недостаток SH2 при наличии АДФ – скорость ограничена.
3. Есть SH2 и АДФ – дыхание очень активно (лимитируется только скоростью транспорта ионов через мембрану).
4. Недостаток АДФ при наличии SH2 – дыхание тормозится (состояние дыхательного контроля).
5. Недостаток кислорода, при наличии SH2 и АДФ – состояние анаэробиоза.
Митохондрии в покоящейся клетке находятся в состоянии 4, при котором скорость дыхания определяется количеством АДФ. Во время усиленной работы могут пребывать в состоянии 3 (исчерпываются возможности дыхательной цепи) или 5 (недостаток кислорода) – гипоксии.
Ингибиторы ЦТД – это лекарственные препараты, которые блокируют перенос электронов по ЦТД. К ним относят: барбитураты (амитал), которые блокируют транспорт электронов через I комплекс дыхательной цепи, антибиотик антимицин блокирует окисление цитохрома b; монооксид углерода и цианиды ингибируют цитохромооксидазу и блокируют транспорт электронов на кислород.
Ингибиторы окислительного фосфорилирования (олигомицин) – это вещества, которые блокируют транспорт Н по протонному каналу АТФ-синтазы.
Разобщители окислительного фосфорилирования (ионофоры) – это вещества, которые подавляют окислительное фосфорилирование, не влияя при этом на процесс переноса электронов по ЦТД. Механизм действия разобщителей сводится к тому, что они являются жирорастворимыми (липофильными) веществами и обладают способностью связывать протоны и переносить их через внутреннюю мембрану митохондрий в матрикс, минуя протонный канал АТФ-синтазы. Выделяющаяся при этом энергия рассеивается в виде тепла.
Искусственные разобщители – динитрофенол, производные витамина К (дикумарол), некоторые антибиотики (валиномицин).
Естественные разобщители – продукты перекисного окисления липидов, жирные кислоты с длинной цепью, большие дозы йодсодержащих гормонов щитовидной железы, белки термогенины.
На разобщении дыхания и фосфорилирования базируется терморегуляторная функция тканевого дыхания. Митохондрии бурой жировой ткани продуцируют больше тепла, так как присутствующий в них белок термогенин разобщает окисление и фосфорилировние. Это имеет важное значение в поддержании температуры тела новорожденных.
Глава 11. Типы окисления. Антиоксидантные системы
Все реакции с участием кислорода, протекающие в живом организме, называются биологическим окислением. Почти во всех клетках около 90 % потребляемого кислорода восстанавливается в цепи тканевого дыхания с участием цитохромоксидазы (окисление, сопряженное с фосфорилированием АТФ, выполняет энергетическую функцию). Однако в некоторых тканях содержатся ферменты, катализирующие окислительно-восстановительные реакции, в которых атомы кислорода включаются непосредственно в молекулу субстрата (свободное окисление, выполняет пластическую функцию). Хотя в таких специализированных реакциях потребляется лишь небольшая часть кислорода, поглощаемого клетками, эти реакции очень важны для организма.
Выделяют четыре типа реакций с участием кислорода (табл 11.1.).
Таблица 11.1. Типы окисления
| Тип окисления | Ферменты | Основные продукты реакции |
|---|---|---|
| оксидазный | Оксидазы | S + Н2О |
| пероксидазный | ФАД-зависимые оксидазы | S + Н2О2 |
| диоксигеназный | Диоксигеназы | SO2 |
| монооксигеназный | Монооксигеназы (гидроксилазы) | SOH + H2O |
Оксидазный тип окисления
Этот путь окисления осуществляется в процессе функционирования ЦТД. Терминальный фермент ЦТД, переносящий электроны непосредственно на кислород – цитохромоксидаза. Это основной путь потребления кислорода в организме. Он выполняет энергетическую функцию.
Пероксидазный тип окисления
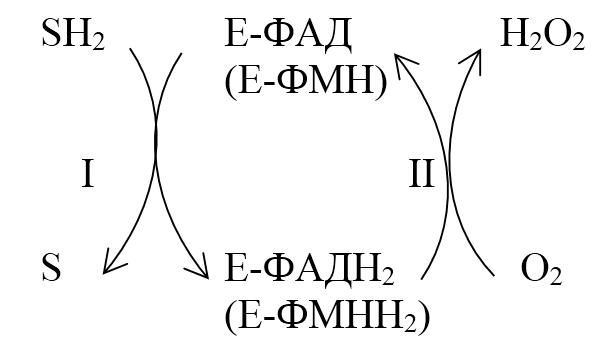
Окисление субстрата путем дегидрирования. Два атома водорода переносятся на молекулу кислорода с образованием перекиси:
ФАД-зависимая оксидаза
SН2 + О2 → S + Н2О2
В этой реакции энергия окисления выделяется в виде тепла. Реакции этого типа катализируют ФАД-зависимые оксидазы (аэробные дегидрогеназы), содержащие в качестве простетической группы ФАД или ФМН. В клетке около 80 % этих ферментов сосредоточено в пероксисомах. Пероксидазный путь окисления активно протекает в лейкоцитах, макрофагах и других фагоцитирующих клетках. Образующийся пероксид водорода Н2О2 – сильный окислитель, обезвреживающий патогенные бактерии (защитная функция).
Реакция пероксидазного окисления протекает в 2 стадии:
1. Анаэробная - происходит дегидрирование восстановленного субстрата SH2, при этом протоны и электроны переносятся на ФАД (ФАД + 2Н → ФАДН2).
2. Аэробная - происходит окисление фермента (ФАДН2 → ФАД) кислородом (самопроизвольный процесс).
Биологическая роль оксидазного типа окисления:
1. защитная функция – в лейкоцитах и других фагоцитирующих клетках;
2. катаболизм биогенных аминов (фермент – моноаминооксидаза);
3. метаболизм аминокислот (ферменты – оксидазы D- и L-аминокислот);
4. катаболизм пуринов (фермент – ксантиноксидаза);
5. катаболизм глюкозы в растительных клетках (фермент – глюкозооксидаза).
Диоксигеназный тип окисления
В процессе диоксигеназного окисления в молекулу субстрата включаются оба атома кислорода:
Диоксигеназа
S + O2 → SO2
Диоксигеназы катализируют разрыв двойной связи в ароматическом кольце. Например: гомогентизатоксидаза катализирует расщепление ароматического кольца гомогентизиновой кислоты с образованием малеилацетоацетата.
Монооксигеназный тип окисления
Монооксигеназы (гидроксилазы) катализируют включение в субстрат одного атома молекулы кислорода. Другой атом кислорода восстанавливается до воды. Для работы монооксигеназной системы необходим кроме неполярного субстрата (SH) донор атомов водорода – косубстрат (НАДФН+Н, ФАДН2, аскорбиновая кислота):
Гидроксил
SH + НАДФН+Н+ О2 → SОН + НАДФ + Н2О
Монооксигеназные реакции необходимы для:
1. специфических превращений аминокислот, например, для синтеза тирозина из фенилаланина (фермент – фенилаланингидроксилаза);
2. синтеза холестерола, желчных кислот в печени; стероидных гормонов в коре надпочечников, яичниках, плаценте, семенниках; витамина D3 в почках;
3. обезвреживания чужеродных веществ (ксенобиотиков) в печени.
Ферменты монооксигеназного пути окисления локализованы в мембранах эндоплазматического ретикулума (при гомогенизации тканей эти мембраны превращаются в микросомы – мембранные пузырьки). Поэтому монооксигеназный путь окисления называют микросомальным окислением.
Микросомальное окисление представляет короткую электронтранспортную цепь, включающую НАДФ, ФАД, ФМН, цитохром Р450.
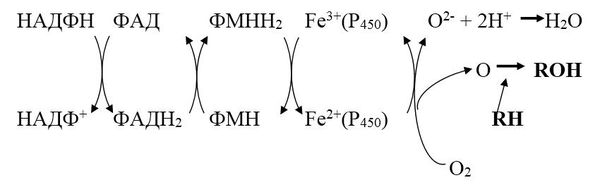
Микросомальная система включает два фермента: цитохром Р450 и НАДФН-цитохром-Р450-редуктазу.
НАДФН-цитохром Р450 – редуктаза – флавопротеин, в качестве простетической группы содержит два кофермента ФАД и ФМН.
Цитохром Р450 – гемопротеин, содержит простетическую группу гем и участки связывания для кислорода и субстрата. Восстановленный цитохром Р450 имеет максимум поглощения при 450 нм. Выполняет две функции: связывание окисляемого субстрата и активация молекулярного кислорода.