Всего за 149 руб. Купить полную версию

Рис. 8. Человек с острова Флорес (Homo floresiensis) обитал на одном из Малых Зондских островов в Индонезии 74–12 тыс. лет назад, когда всей планетой уже владел биологический вид Homo sapiens. "Флоресиец" представлял собой позднюю карликовую форму гейдельбергского человека и достигал всего 106 см роста (судя по женскому экземпляру). Он не являлся ни сказочным хоббитом ("чертенком"), ни позднейшим питекантропом, а сделался крохой подобно другим карликовым островным млекопитающим (таким, как карликовые мамонты о-ва Врангеля или вымершие карликовые слоны о-ва Мальта) [23а]
Австралоиды – монголоиды – 83,9 ± 3,1 тыс. лет назад (Н). Азия: Люджянг, Китай, 67 тыс. лет назад [214]; Япония, 70–50 тыс. лет назад [160]. Австралазия: Ниах грейт кейв, Саравак, Калимантан, Малайзия, св. 41500 ± 100 лет назад; Хуон, Папуа – Новая Гвинея, 53–45 тыс. лет назад [382]; Матенкупкум, Новая Ирландия, северная Меланезия, 32700 ± 1550 лет назад [135, с. 709]; Малакунанджа II, западный откос плато Арнемленд, Австралия, 61 ± 13–45 ± 9, 52 ± 11 тыс. лет назад [648]. Америка: Монте Верде, Чили, 33370 ± 530 лет назад; Тока ду Бокейрану да Педра Фурада, Бразилия, св. 32 тыс. лет назад [385; 500, с. 708].
Представители рода Homo отличались строением тела, приспособленным к прямохождению. Они имели относительно короткие руки и относительно длинные ноги, как у двуногих динозавров. В отличие от прочих млекопитающих, наши предки обладали не слишком различающимися резцами, клыками и коренными зубами, т. е. были равнозубыми, как динозавры, что объясняется их ориентацией на высокобиопродуктивную среду, когда не требуется тщательно утилизировать пищу. Сходство усугублялось тем, что некоторые динозавры (ранние карнозавры и целурозавры) располагали хватательными верхними конечностями с отстоящим внутренним пальцем, аналогом нашего большого [25, с. 45–46, 49]. Однако, несмотря на подходящую физическую организацию, мы все еще не обладаем твердыми наследственными навыками прямохождения. Это доказывается судьбами homo ferus (детьми, выращенными животными), которые без человеческого воспитания к прямохождению не способны (ползают на четвереньках) [76, с. 47–49, 183–186; 576].
Следует добавить, что, хотя люди преодолевают стометровку не быстрее крохотной, но четвероногой крысы (ок. 10 с в обоих случаях), они все же кое в чем продвинулись как выносливые бегуны. Предки рода Homo были приспособлены к выносливому бегу гораздо хуже [207], что отвечает большей неотеничности. Способность к выносливому бегу появилась лишь у Kenyanthropus rudolfensis или Homo ergaster в результате акселерации (противоположной неотении, см. ниже). Человек современного вида перенес до трех акселераций, а потому оказался способен к марафону, хотя все еще десятикратно уступает в этом четвероногим гиенам или волкам.
Представители остальных родов гоминин, будучи тоже прямоходящими, сохраняли обезьянье строение тела. Они имели относительно длинные, питекоидные руки (Australopithecus habilis, Australopithecus afarensis, Ardipithecus ramidus [36; 145, с. 556; 442; 443]), а также обезьянье строение вестибулярного аппарата, существенного для прямохождения (Australopithecus habilis, Paranthropus robustus [720]). Иными словами, история прямохождения у гоминин, от самых ранних до современных, постепенна и не закончена до сих пор.
Прямохождение возникло именно у наших обезьяноподобных предков благодаря эволюционной отсталости приматов, делающей их податливыми для фундаментальных перемен. Среди млекопитающих эволюционно продвинуты не они, а мышевидные грызуны [82, с. 97]. Энергетически экономная двуногость явилась откликом на подъем биопродуктивности среды в связи с потеплением, когда подвижный поиск пищи стал неактуальным, раз она обильна и скучена в пространстве. Подобная кормовая база спровоцировала у гоминин "мутацию прожорливости". Они стали потреблять примерно в 3,8 раза больше пищевой энергии на единицу массы тела, нежели все остальные высшие млекопитающие. У последних соответствующий показатель (постоянная Рубнера) составляет 191 600 ккал на кг массы тела в течение жизни, тогда как у людей это 725 800 ккал на кг телесной массы [6, с. 73, 89; 661; 662]. Таким образом, метаболизм (обмен веществ) у людей по сравнению с другими высшими млекопитающими завышен в 3,8 раза.
Неотения и акселерация
Вынужденные перерабатывать почти вчетверо больше энергии, чем равновеликие млекопитающие, гоминины стали пропорционально дольше жить [323] и, соответственно, формироваться. Их индивидуальное развитие затормозилось, растянулось во времени и привело к сохранению инфантильных (детских) и даже эмбриональных черт во взрослом состоянии, что называется неотенией (от греч. νεότης – юность) (рис. 9) [10; 12, с. 17–19; 402; 577; 584, с. 13–44]. Аналогичное явление (педоморфоз) известно у земноводного аксолотля [564]. У гоминин неотения выразилась в удержании эмбриональной обезволошенности тела и невыраженности лицевого скелета, в силу чего мозговой скелет, купол черепа, непропорционально вздулся. Это не связано с интеллектуальными нагрузками: у испытывающего их последние две с половиной тыс. лет сапиенса мозг ничуть не изменился. Известно, что вздутие купола черепа началось у Australopithecus afarensis до того, как усложнилась кортикальная организация его мозга (организация его мозговой коры) [315]. Проще сказать, у гоминин обезьяний по сути мозг механически разросся в силу неотении, не усложняясь как орган интеллекта. Отметим, что старший современник "афарца", Australopithecus africanus из Таунга в ЮАР, по стадиям прорезывания зубов напоминал не ребенка 5–7 лет (как следовало бы из его вероятного возраста), а детеныша человекообразной обезьяны 3–4 лет [257, с. 626] в силу того, что фенотипическая (телесная) неотения отставала у гоминин от генотипической. Аналогичным образом наша генетическая предрасположенность к прямохождению еще не гарантирует, что человеческий ребенок усвоит двуногость автоматически.
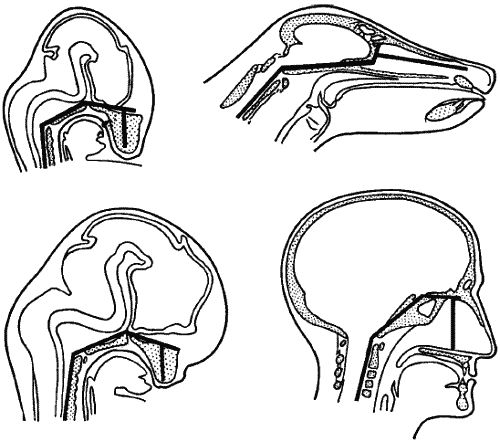
Рис. 9. Человек и его предки гоминины возникли в результате эволюции путем неотении, когда организм во взрослом состоянии сохраняет детские и даже эмбриональные (зародышевые) особенности. У человека к ним относятся обезволошенность тела (эмбрионы человека и многих других млекопитающих безволосы), а также вздутый купол черепа. У эмбрионов человека и прочих высших млекопитающих он вздут, однако после рождения у животных сильно разрастается морда, и мозговой участок черепа выглядит убогим; у людей этого не происходит: лицо остается крохотным, а потому мозг кажется обширным. По общему (ошибочному) убеждению это объясняется большим интеллектом людей. Однако человеческий ум существенно развивался только у Homo sapiens, да и то лишь последние 2,5 тыс. лет. При этом мозг человека современного типа не изменился ни на йоту. Вдобавок наш мозг активно загружен не более чем на 10%. Здесь мы видим эмбриональные и зрелые формы черепа у собаки и у человека. Оба эмбриональных абриса похожи, но лишь у человека зародышевые очертания сохраняются во взрослом состоянии [12, с. 19]
Под влиянием неотении, замедлившей индивидуальное развитие наших предков, у женщин и самок гоминин месячный цикл растянулся, так что эпизоды способности к зачатию как бы слились, в результате чего эструс (аналог течки у самок животных) размазался и стерся, хотя у женщин сохраняется несколько ежемесячных дней, когда они повышенно восприимчивы к ухаживаниям.
Самки гоминин стали существенно дольше, чем равновеликие млекопитающие, вынашивать потомство, что вызывало накопление ими заметных жировых запасов, необходимых для питания плода [107; 402; 583; 628]. Начиная с питекантропа, изменились сроки прорезывания зубов у молоди наших предков [280]. На протяжении своей эволюции гоминины все заметнее уподоблялись эмбрионам, что выразилось у них в грацилизации, т. е. в утончении костей скелета и приобретении изящества. Ныне это явление усиливается благодаря цивилизованному образу жизни: он вызывает последствия, напоминающие эффекты одомашнивания животных, которое изнеживает, инфантилизирует наших питомцев [490, с. 357–358].
Неотеническое увеличение продолжительности жизни у гоминин имело эволюционные последствия. Темпы эволюции зависят от скорости смены поколений. Поэтому замедление онтогенеза влечет за собой торможение филогенеза вида. У гоминин он в 3,8 раза замедлен против равновеликих млекопитающих. Поэтому, сравнивая их молекулярную эволюцию с нашей, мы должны умножать хронометраж на своего рода "неотенический коэффициент" 1,894, чтобы привести к среднему знаменателю нормально текущую эволюцию с нашей, вчетверо заниженной. Как можно видеть из вышеприведенного хронометрированного списка гоминин, подобная коррекция молекулярно-генетических дат (М) при помощи "неотенического коэффициента" дает исправленные датировки (Н), совместимые с палеоантропологическими данными. Заторможенность молекулярного мутагенеза у человека как таковая выявлена эмпирически [506], однако она не дотягивает до заторможенности человеческого онтогенеза, что объясняется сложностью расчетов абсолютных количественных показателей для мира млекопитающих, где они зависят от размеров, физиологии и других особенностей животных. Так, продолжительности жизни кошки, человека и кита относятся примерно как 1:10:20, в то время как массы их тел относятся друг к другу как 1:10:1500, откуда видно, что рост продолжительности жизни у млекопитающих заметно отстает от увеличения их размеров.