Недостаточное поступление желчи в кишечник называется гипохолией, полное прекращение ее поступления – ахолией. Эти явления возможны при закупорке или сдавлении общего желчного протока, при нарушении желчеобразовательной функции печени. При ахолии резко страдает переваривание и всасывание жира, так как липаза поджелудочного сока в отсутствие желчи малоактивна, а жиры не эмульгированы и контакт их с липолитическим ферментом затруднен. При недостатке желчи страдает всасывание жирных кислот, холестерина, жирорастворимых витаминов. Вследствие недостаточного переваривания и всасывания жира развивается стеаторея – избыточное содержание жира в испражнениях. С калом при этом выводится до 70–80 % жира. В кишечнике нерасщепленный жир обволакивает пищевые массы и затрудняет действие на них панкреатических протеолитических ферментов и α-амилазы. Гидролитическая активность ферментов при недостатке желчи падает, так как желчь принимает участие в нейтрализации кислого желудочного содержимого и способствует поддержанию в двенадцатиперстной кишке щелочной среды, являющейся оптимальной для ферментов дуоденального сока.
Наконец, желчные кислоты стимулируют сорбционные свойства кишечного эпителия, поэтому при их дефиците уменьшается адсорбция кишечными клетками ферментов из химуса и нарушается мембранное пищеварение. Таким образом, при гипо– и ахолии нарушаются также начальные и промежуточные этапы переваривания белков и углеводов в тонкой кишке.
Вместе с неусвоенными жирами из кишечника выводятся и жирорастворимые витамины. Развиваются гиповитаминозы, в частности, нередко понижается свертываемость крови в связи с дефицитом витамина К.
При гипо– и ахолии ослабляется перистальтика кишечника, что в сочетании с выпадением бактерицидного действия желчи ведет к усилению в кишечнике процессов брожения и гниения, к метеоризму – развивается дисбактериоз кишечника. В результате наступает интоксикация организма.
Рефлекторные влияния на желчевыделительный аппарат осуществляются условно– и безусловно-рефлекторно с участием многочисленных рефлексогенных зон, в том числе рецепторов полости рта, желудка и двенадцатиперстной кишки. Велика роль гуморальной регуляции желчевыделения. Большую роль в качестве гуморального стимулятора желчевыделения играет холецистокинин, вызывающий сокращение желчного пузыря. Сокращения желчного пузыря, хотя и слабые, вызывают также гастрин, секретин, бомбезин. Тормозят сокращения желчного пузыря глюкагон, кальцитонин, антихолецистокинин, ВИП,ПП.
Глава 3 Базисные сведения о важнейших видах обмена веществ
Метаболизм основных видов нутриентов в норме и при патологии определяет потребности организма в них и пути их введения. Поэтому знания о процессах ассимиляции различных питательных веществ являются основой диетической терапии.
Белки
Белки – сложные азотсодержащие биополимеры, мономерами которых служат аминокислоты (органические соединения, содержащие карбоксильную и аминогруппы). Их биологическая роль многообразна. Белки выполняют в организме пластические, каталитические, гормональные, транспортные и другие функции, а также обеспечивают специфичность. Значение белкового компонента питания заключается, прежде всего, в том, что он служит источником аминокислот.
Аминокислоты делятся на эссенциальные и неэссенциальные в зависимости от того, возможно ли их образование в организме из предшественников. К незаменимым аминокислотам относятся гистидин, лейцин, изолейцин, лизин, метионин, фенилаланин, триптофан и валин, а также цистеин и тирозин, синтезируемые соответственно из метионина и фенилаланина. 9 заменимых аминокислот (аланин, аргинин, аспаргиновая и глутаминовая кислоты, глутамин, глицин, пролин и серии) могут отсутствовать в рационе, так как способны образовываться из других веществ. В организме также существуют аминокислоты, которые продуцируются путем модификации боковых цепей вышеперечисленных (например, компонент коллагена – гидроксипролин – и сократительных белков мышц – 3-метилгистидин).
Большинство аминокислот имеют изомеры (D– и L-формы), из которых только L-формы входят в состав белков человеческого организма. D-формы могут участвовать в метаболизме, превращаясь в L-формы, однако утилизируются гораздо менее эффективно.
По химическому строению аминокислоты делятся на двухосновные, двухкислотные и нейтральные с алифатическими и ароматическими боковыми цепями, что имеет важное значение для их транспорта, поскольку каждый класс аминокислот обладает специфическими переносчиками. Аминокислоты с аналогичным строением обычно вступают в сложные, часто конкурентные взаимоотношения.
Так, ароматические аминокислоты (фенилаланин, тирозин и триптофан) близкородственны между собой. Хотя фенилаланин является незаменимой, а тирозин синтезируемой из него заменимой аминокислотой, наличие тирозина в рационе как будто бы "сберегает" фенилаланин. Если фенилаланина недостаточно, или его метаболизм нарушен (например, при дефиците витамина С) – тирозин становится незаменимой аминокислотой. Подобные взаимоотношения характерны и для серосодержащих аминокислот: незаменимой – метионина, и образующегося из него цистеина.
Приведем другой пример. Триптофан в ходе превращений, для которых необходим витамин В6 (пиридоксин), включается в структуру НАД и НАДФ, то есть дублирует роль ниацина. Приблизительно половина обычной потребности в ниацине удовлетворяется за счет триптофана: 1 мг ниацина пищи эквивалентен 60 мг триптофана. Поэтому состояние пеллагры может развиваться не только при недостатке витамина РР в рационе, но и при нехватке триптофана или нарушении его обмена, в том числе вследствие дефицита пиридоксина.
Аминокислоты также делятся на глюкогенные и кетогенные, в зависимости от того, могут ли они при определенных условиях становиться предшественниками глюкозы или кетоновых тел (табл. 3.1).
Таблица 3.1 Классификация аминокислот
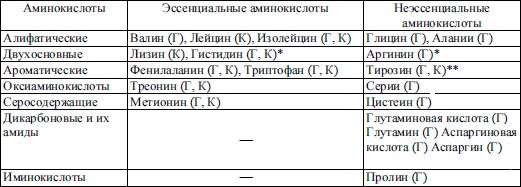
Примечания: Г – глюкогенные, К – кетогенные аминокислоты; * – гистидин незаменим у детей до года; ** – "условно-незаменимые" аминокислоты (могут синтезироваться из фенилаланина и метионина).
Поступление азотсодержащих веществ с пищей происходит в основном за счет белка и, в менее значимых количествах, свободных аминокислот и других соединений. В животной пище основное количество азота содержится в виде белка. В продуктах растительного происхождения большая часть азота представлена небелковыми соединениями, а также содержится множество аминокислот, которые не встречаются в организме человека и зачастую не могут метаболизироваться им.
Человек не нуждается в поступлении с пищей нуклеиновых кислот. Пуриновые и пиримидиновые основания синтезируются в печени из аминокислот, а избыток этих оснований, поступивший с пищей, выводится в виде мочевой кислоты. В синтезе пиримидиновых колец принимает участие витамин В12, для образования пуриновых структур необходима фолиевая кислота. Именно поэтому дефицит этих нутриентов отражается, прежде всего, на органе с высоким уровнем пролиферации, где идет наиболее интенсивный синтез нуклеиновых кислот – кроветворной ткани.
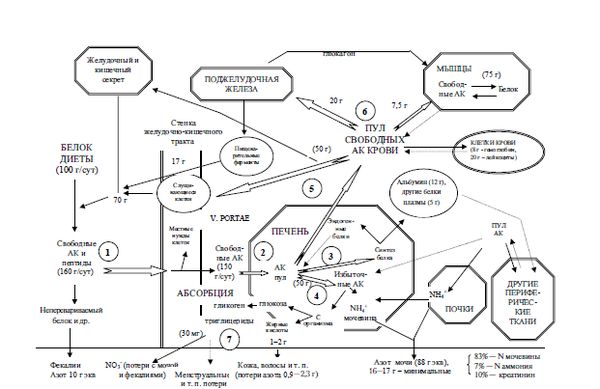
Обычный (но не оптимальный) ежедневный прием белка у среднестатистического человека составляет приблизительно 100 г. К ним присоединяется примерно 70 г белка, секретируемого в полость желудочно-кишечного тракта. Из этого количества абсорбируется около 160 г. Самим организмом в сутки синтезируется в среднем 240–250 г белка. Такая разница между поступлением и эндогенным преобразованием свидетельствует об активности процессов ресинтеза (рис. 3.1).
Для здорового человека характерно состояние азотного равновесия, когда потери белка (с мочой, калом, эпидермисом и т. п.) соответствуют его количеству, поступившему с пищей. При преобладании катаболических процессов возникает отрицательный азотный баланс, который характерен для низкого потребления азотсодержащих веществ (низкобелковые рационы, голодание, нарушение абсорбции белка) и многих патологических процессов, вызывающих интенсификацию распада (опухоли, ожоговая болезнь и т. п.). При доминировании синтетических процессов количество вводимого азота преобладает над его выведением и возникает положительный азотный баланс, характерный для детей, беременных женщин и реконвалесцентов после тяжелых заболеваний.
После прохождения энтерального барьера белки поступают в кровь в виде свободных аминокислот. Следует отметить, что клетки слизистой оболочки желудочно-кишечного тракта могут метаболизировать некоторые аминокислоты (в том числе глутаминовую кислоту и аспаргиновую кислоту в аланин). Способность энтероцитов видоизменять эти аминокислоты, возможно, позволяет избежать токсического эффекта при их избыточном введении.
Аминокислоты, как поступившие в кровь при переваривании белка, так и синтезированные в клетках, в крови образуют постоянно обновляющийся свободный пул аминокислот, который составляет около 100 г.
75 % аминокислот, находящихся в системной циркуляции, представлены аминокислотами с ветвящимися цепями (лейцином, изолейцином и валином). Из мышечной ткани в кровоток выделяются аланин, который является основным предшественником синтеза глюкозы, и глутамин.