Всего за 259.9 руб. Купить полную версию
С моей точки зрения, структура ДНК и модель ее репликации, которую Уотсон и Крик описали в своей второй классической статье как непосредственное следствие структуры (Watson and Crick, 1953a), являются важнейшим фундаментальным открытием в изучении эволюции со времени публикации "Происхождения видов…". По сути, Уотсон и Крик вывели из структуры ДНК биологическое воплощение общего принципа цифрового хранения, кодирования и передачи информации. Система биологической передачи информации, которую выявили их исследования, может рассматриваться как расширение принципа машины Тьюринга, сначала через правила комплементарности нуклеотидных оснований (в процессах репликации и транскрипции), a затем в процессе трансляции, через генетический код (см. рис. 2–1). По сути, пусть и не в историческом смысле, эти открытия вытеснили концепцию Дарвина, в том смысле, что вся дарвиновская схема эволюции является прямым следствием механизма репликации ДНК. Для всех известных форм жизни биологическая передача цифровой информации влечет за собой исполнение следующих простых фундаментальных принципов.
Генетический материал любого организма состоит из линейной последовательности символов, четырех оснований нуклеиновых кислот, которая, прямо или косвенно, кодирует всю информацию, необходимую для построения организма.
Репликация генетического материала, являющегося механической основой наследственности, осуществляется на основе принципа однозначного комплементарного соответствия между A и T(U), и G и С. (Так называемые правила Чаргафа, по имени их первооткрывателя, австрийского, a затем американского химика Эрвина Чаргаффа.)
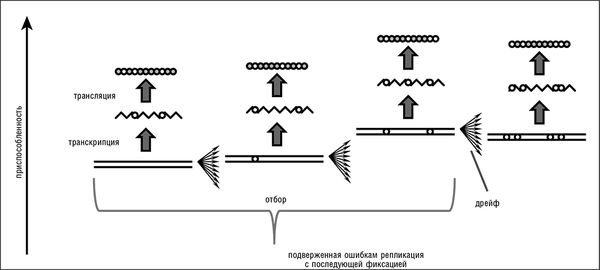
Рис. 2–1. Передача информации в биологических системах и превращение естественного отбора и генетического дрейфа в эпифеномены репликации. Белые круги на схеме обозначают изменения относительно оригинальной последовательности.
Уотсон и Крик описали эти ключевые принципы генетических систем в двух своих статьях, вышедших в 1953 году. Дальнейшие исследования добавили два очень важных аспекта:
1. Принцип комплементарности используется не только во время репликации, но и во время транскрипции ДНК во все виды РНК и во время трансляции мРНК в белок с помощью адапторных молекул тРНК.
2. Те же самые принципы цифровой репликации и декодирования применимы и для генетических систем, в которых генетический материал отличается от двойной спирали ДНК, изначально описанной Уотсоном и Криком, и состоит из РНК или односпиральной ДНК (например, у многих вирусов).
Теория информации твердо стоит на том, что передача информации абсолютно без ошибок невозможна в принципе. В реальности вероятность ошибки в любом конечном сообщении может быть сведена к минимуму, но любое снижение уровня ошибок при передаче информации возможно только за счет затраты энергии. Эта связь непосредственно следует из законов термодинамики. Центральный принцип эволюции может быть сформулирован следующим образом:
Репликация цифровых носителей информации неизбежно подвержена ошибкам, что влечет за собой эволюцию этих носителей путем естественного отбора и случайного дрейфа генов при условии, что уровень ошибок репликации ниже катастрофического порога, имеющего порядок величины от одной до десяти ошибок на геном за один цикл репликации.
Назовем это обобщение принципом подверженной ошибкам репликации (ПОР). Этот принцип становится самоочевидным, как только мы осознаем существование и основной механизм репликации. Он был впервые описан математически в теории Манфреда Эйгена (Eigen, 1971), который также ввел понятие концепции порога ошибки (Biebricher and Eigen, 2005) – эта теория и ее применение будут рассматриваться далее в главе 12. ПОР основывается на следующих двух предположениях, которые могут показаться очевидными, но заслуживают тем не менее особого упоминания:
1. Ошибки репликации наследуются (проходят через циклы репликации).
2. Существует обратная связь между генотипом и фенотипом: некоторые ошибки репликации влияют на эффективность и точность репликации как отрицательно, так и положительно.
Это и отличает биологические репликаторы с их "неограниченной наследуемостью" от репликаторов с "ограниченной наследуемостью", таких как кристаллы или ряд химических циклов, которые реплицируются, но не передают накопленные дефекты последующим поколениям (Szathmary, 2000). Упрощенно говоря, разница в том, что в нуклеиновых кислотах замена одного нуклеотида на другой влияет только на передаваемую информацию, а не на физические или химические свойства носителя информации (во всяком случае, не значительно), как в случае небиологических систем.
В принципе должен существовать и нижний порог частоты ошибок репликации. Очевидно, что если математическое ожидание числа ошибок на цикл репликации стремится к нулю, то разнообразие становится недостаточным для эволюционного процесса. Однако необходимо заметить, что уровень ошибок репликации нуклеиновых кислот, определяемый эмпирически, в отсутствие сложных корректирующих механизмов (как, например, в РНК вирусов), не намного ниже верхнего, катастрофического порога. Таким образом, необходимым условием эволюции является достаточно низкий (а не достаточно высокий) уровень ошибок репликации. Вопрос о том, в какой степени оптимизируется фактическая точность передачи информации в биологической системе (другими словами, эволюционирует ли эволюция), весьма сложен, интересен и широко обсуждается. Мы рассмотрим его подробнее в главе 9.
Несмотря на то что вся естественно эволюционировавшая жизнь основана на репликации нуклеиновых кислот, ПОР не зависит от физической природы репликаторов, как можно видеть на примере эволюции компьютерных вирусов и различных компьютерных моделей эволюции искусственной жизни (Lenski et al., 2003). Тем не менее вопрос о том, необходим ли цифровой код для эволюции или эволюция может происходить и в аналоговых системах, весьма интересен и до сих пор остается открытым.
В главе 1 был затронут вопрос o псевдотавтологичной природе естественного отбора. По сути, ПОР действительно в значительной мере тривиализирует естественный отбор и генетический дрейф (два фундаментальных принципа эволюции), как бы отбирая у них статус независимых феноменов и низводя их до эпифеноменов ПОР. Это ни в коей мере не умаляет достижения Дарвина, Райта и других выдающихся эволюционных биологов и не уменьшает важность концепций естественного отбора и генетического дрейфа для описания эволюционных процессов на абстрактном уровне. Тем не менее открытие репликации с контролируемым уровнем ошибок обнаруживает более фундаментальные принципы, которые лежат в основе классических положений эволюционной биологии.