Когда говорят о структуре мезозойского сообщества наземных позвоночных, сразу замечают, что крупный размерный класс (Э. Олсон назвал его "доминантным сообществом") в нем был полностью сформирован архозаврами: и фитофаги, и хищники в нем представлены сперва текодонтами, потом динозаврами. Реже обращают внимание на другое обстоятельство: малый размерный класс ("субдоминантное сообщество") оказался для архозавров почти закрытым - ровно в той же степени, как крупный - для тероморфов. Среди малоразмерных существ (менее 1 м) доминировали териодонты (и их прямые потомки - млекопитающие), а на вторых ролях выступали низшие диапсиды - ящерицы и клювоголовые (ныне от этой группы уцелела лишь гаттерия); питались они насекомыми и, реже, друг другом (фитофагии в малом размерном классе не было вовсе). Итак, высшие завроморфы (динозавры) и высшие тероморфы (маммалии) с самого начала сформировали "параллельные миры", практически не взаимодействовавшие между собой.
Очень интересно сопоставить структуру сообщества наземных, вторично-водных и летающих позвоночных. Если на суше господствовали амниоты с самым активным метаболизмом - тероморфы и архозавры (последние имели четырехкамерное сердце и, как мы увидим дальше, по-своему решили проблему гомойотермии), то водная среда оказалась вотчиной рептилий с низким уровнем обмена и белой мускулатурой, ориентированной на бескислородные процессы. Для архозавров она была малоподходящей (водный образ жизни ведут лишь самые архаичные их представители, фитозавры и крокодилы), зато здесь процвели низшие диапсиды (эвриапсиды) и анапсиды: ихтиозавры, плезиозавры, плакодонты, морские черепахи. Нет такой группы диапсид, которая не "делегировала" бы своих представителей в воду - были даже водные клювоголовые, плеврозавры.
Все водные рептилии - хищники (среди них не было не только фитофагов, но даже и планктоноядов вроде усатых китов или китовой акулы), и все крупные - заметно крупнее своих сухопутных предков (если таковые известны). Весьма показательны в этом последнем отношении "ящерицы" (в широком смысле), на суше практически никогда не выходившие из малого размерного класса, но породившие гигантских морских мозазавров и пресноводных хампсозавров (жизненная форма "крокодил"). Начиная с триаса водные рептилии оккупировали верхние трофические этажи морских экосистем, полностью вытеснив оттуда и рыб, и головоногих - эти предыдущие "владыки морей" служили им основной пищей. Ситуация изменилась лишь в начале мела, когда появились акулы современного типа; возможно, именно они конкурентно вытеснили самых "рыбообразных" из рептилий - ихтиозавров.
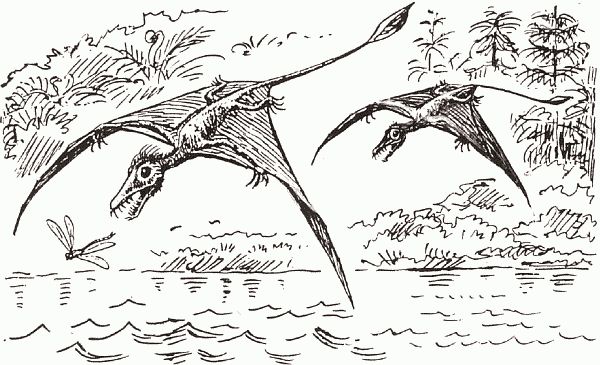
С освоением воздуха картина, как и следовало ожидать, была обратная (по сравнению с морем): здесь преуспели именно высшие архозавры, имевшие самый активный метаболизм, - птицы и птерозавры, которые тоже почти наверняка были теплокровными. Так, мелкий птерозавр Sordes pilosus (что в переводе с латыни означает "нечисть волосатая") был покрыт чем-то вроде волосяного покрова не вполне ясной природы; трудно представить себе, зачем он мог понадобиться, если не для теплоизоляции. В пользу теплокровности птерозавров может косвенно свидетельствовать и строение их костей, полых и несущих перфорации - как у птиц, у которых кости пронизаны воздушными мешками (обеспечивающими бесперебойность дыхания во время машущего полета). Если дыхательная система в этих двух группах построена аналогичным образом, то и интенсивность обмена у птерозавров должна быть очень высока.

Самое интересное, что в триасе (и даже в конце перми) низшие диапсиды ("ящерицы") совершили целый ряд попыток завоевания воздуха, некоторые из которых кажутся крайне экзотичными (рис. 47, а–в), однако это были варианты планирующего полета: на активный, машуший, полет у этих рептилий явно не хватает энергетики. Появившиеся в конце триаса воздушные архозавры (птерозавры) разом пресекли эти неуклюжие потуги низших диапсид. Первыми птерозаврами были хвостатые рамфоринхи, размером от воробья до вороны; в дальнейшем птерозавры становились все более крупными (например, птеродактили были не меньше орла, а у их верхнемеловых представителей, птеранодонов, размах крыльев достигал 8 м - это самые большие летающие существа всех времен) и наверняка перешли к парящему полету, вроде современных кондоров. Исчезновение в конце юры мелких высокоманевренных рамфоринхов (хвост летающему существу нужен как противовес для разворотов с малым радиусом) с машущим полетом, после чего остались лишь крупноразмерные "живые планеры" - птеродактили, судя по всему, было напрямую связано с появлением в это время птиц. В воздушном сообществе между птерозаврами и птицами происходит такое же размежевание по размерным классам, как и в наземном - между динозаврами и маммалиями. Рамфоринхи (судя по содержимому их желудков) были частью насекомоядными, частью рыбоядными - что-то вроде современных зимородков (рис. 48). В дальнейшем рыбоядность стала для птерозавров "генеральной линией" - недаром почти все их скелеты найдены в мелководных морских отложениях.
Итак, в море господствуют крупные рептилии из числа низших завроморфов с "холодным" метаболизмом, в воздухе - относительно мелкие и теплокровные высшие архозавры. А что же "владыки суши" - динозавры? Вопрос об их возможной теплокровности дискутировался давно. В последнее время большинство палеонтологов сходятся на том, что интенсивность обмена у динозавров может быть и не достигала птичьей или маммальной, но заметно превышала таковую "обычных" рептилий. Во всяком случае, ранние представления о динозаврах как о массивных малоподвижных существах отошли в прошлое. Ужасные тероподы-велоцирапторы из "Парка юрского периода", напоминающие своим поведением крупных нелетающих птиц, судя по всему, вполне соответствуют реальности.
У динозавров тело было приподнято над землей гораздо больше, чем у любых современных рептилий с их широко расставлеными в стороны конечностями. Об этом свидетельствуют, в числе прочего, их узкие следовые дорожки - как у маммалий и птиц. Иногда указывают на то, что динозавры просто не могли нести многотонное тело на своих не до конца выпрямленных ("раскоряченных" по-рептильному) конечностях - это невозможно: надо, чтобы вес принимали на себя не мышцы, а вертикально поставленные кости (см. выше). Верно. Но все-таки это, пожалуй, не причина, а следствие. Ведь "прямохождение" выработалось уже у самых первых динозавров - небольших существ с комплекцией кенгуру, перед которыми проблема избыточного веса еще не стояла. Скорее наоборот: само происхождение динозавров было связано с общей активизацией обмена веществ. Это открыло им путь к парасагитальности (для нее, как мы помним, необходима продолжительная мышечная активность; следовательно - красная мускулатура, следовательно - аэробный метаболизм и теплокровность, и т.д.), а уже парасагитальность, в свою очередь, позволила наращивать размеры тела аж до завроподных 30 тонн.