
Рисунок 10. Кончик листа папоротника в бирманском янтаре.
Наши знания о растениях, произраставших в Альберте в течение кампанского века позднего мела действительно весьма обширны благодаря совместным усилиям многих палеонтологов, работавших десятилетиями21-27. Теперь в лесах, вероятно, совместно произрастали подокарпусы и смолоносные деревья каури (араукариевые). Количество покрытосеменных растений значительно увеличилось, и в это время некоторые из растений, произрастающих в северных широтах, почти наверняка были листопадными. Вопрос о размерах и габитусе тех древних цветковых растений, и о том, какой вклад они могли вносить в экосистему, открыт для обсуждения. Превратились ли некоторые из них к этому времени в деревья, или они лишь покрывали землю или представляли собой низкорослые кустарники? Как сказали Деннис Браман и Ева Коппельхас о позднемеловых ископаемых покрытосеменных Канады, "если у них был древовидный габитус, можно было бы ожидать находок крупных кусков фоссилизированной древесины. Однако в настоящее время древесина покрытосеменных растений неизвестна"21. Поэтому мы можем лишь строить предположения относительно того, какова была их роль в формировании ярусов этих лесов.Существует множество факторов, которые определяют, где будет расти растение, в том числе температура, доступность воды из-за осадков и испарения, тип почвы, дренирование, освещённость, географическая широта, ландшафт и высота над уровнем моря. На любом из янтароносных участков эти параметры менялись со временем, оказывая влияние на растения, которые могли процветать там непрерывно на протяжении ряда тысячелетий. И параллельно тому, как постепенно изменялись распространение и относительная важность растений различных типов на протяжении всего мелового периода, когда одни семейства, роды и виды

Рисунок 11. Колосок примитивного злака Programinis burmitis в бирманском янтаре17.

Рисунок 12. Группа грибов-дождевиков в бирманском янтаре.
вымирали, а другие превращались в более современные формы, животные, которые использовали их в пищу и как укрытие, также претерпевали изменения.Мы знаем, что виды динозавров появлялись и исчезали. Фактически, за время от 25 до 30 миллионов лет, отделяющее одни отложения янтаря от других, фауна сменялась новыми видами и родами множество раз. Питер Додсон полагает, что один отдельно взятый вид динозавров существовал всего лишь от 1 до 2 миллионов лет, тогда как примерное время существования рода было в среднем около 7,7 миллионов лет28. Это означает, что фауна динозавров, возможно, сменялась целиком с каждой геологической эпохой, или примерно десять раз за весь меловой период29. Из-за высокой скорости смены видового состава, относительно немногочисленных местонахождений ископаемых остатков и всесветного распространения динозавров в течение продолжительного рассматриваемого периода времени для изучения доступны лишь немногие общие тенденции изменений30-32. К сожалению, хотя присутствие ископаемых остатков говорит нам о том, какая группа существовала в определённое время в конкретном месте, отсутствие других организмов не означает того, что в течение данного периода времени их не существовало.В меловой период в северном полушарии произошло постепенное изменение средних размеров тела растительноядных динозавров с гигантских многотонных колоссов, господствовавших когда-то в юрский период, на относительно некрупных и более быстроногих травоядных (рис. 13). Особенно сильно размножились и распространились орнитоподы. Возможно, в раннем мелу самыми известными среди них были игуанодоны, которые были весьма обычны в северном полушарии, а близкородственные орнитоподы бродили в это же время по южным континентам. Массивные анкилозавры, украшенные тяжёлыми спинными костными пластинами, инкрустированными множеством роговых бугров и шипов, и вооружённые булавовидным хвостом, стали заметным компонентом раннемеловой фауны в областях, когда-то составлявших Лавразию, но были редкостью в Африке. Стегозавры, эти снабжённые костными пластинами динозавры со смехотворно крохотными головками и двумя рядами шипов, торчащих в задней части их тел, вымерли или стали редкостью в ряде областей. В тех местообитаниях, где они сумели выжить, их присутствие, похоже, не было достаточно значительным. Мелкие пситтакозавры, примитивные рогатые динозавры с клювами, похожими на клюв попугая, стали успешной группой в Азии. Стада зауропод, крупнейших среди четвероногих травоядных, с несоразмерно маленькими головами, венчающими длинные шеи, телами размером с дом и ногами, подобными телефонным столбам, продолжали сотрясать землю неуклюжими шагами на просторах Южной Америки. Вес этих титанов длиной от 80 до 100 футов оценивается в 50 тонн. Однако в других уголках земного шара их можно было наблюдать всё реже и реже. К позднему мелу в северном полушарии начинают впервые появляться новые группы динозавров. И в это время мелкие пахицефалозавры, многие из которых обладали ярко выраженной толстой крышей черепа, окаймлённой рядами костяных бугров, и разнообразные цератопсы с рогами, костяными воротниками и непропорционально крупными головами присоединились к стадам бронированных анкилозавров и двуногих/четвероногих гадрозавров, которые могли достигать 40 футов в длину и весить более 4 тонн. Как ни странно, но оказывается, что на южных континентах сохранились главным образом раннемеловые типы динозавров.
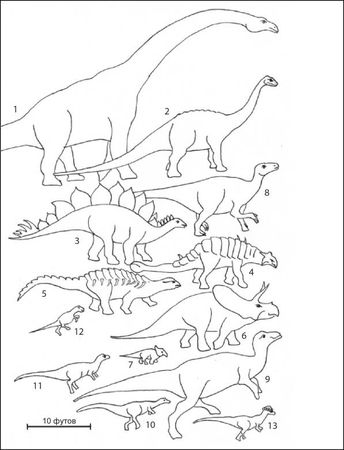
Рисунок 13. Меловые травоядные динозавры, демонстрирующие их относительные размеры. Здесь показаны некоторые из морфотипов травоядных/всеядных динозавров, которые могли жить на одном или нескольких янтароносных участках или вблизи них. Крупнейшими из известных наземных животных были зауроподы. Их примерами были брахиозавриды (1) и титанозавриды (2), а также диплодоциды и цетиозавриды. Другие массивные четвероногие травоядные, но обладающие роговыми "клювами", включают стегозаврид (3), анкилозаврид (4), нодозаврид (5) и цератопсид (6). Группой мелких четвероногих динозавров были протоцератопсиды (7). Орнитоподы, размер которых варьировал от очень большого до сравнительно мелкого, ходили на двух, а иногда на четырёх ногах и также обладали клювовидными образованиями. Они включали игуанодонтид (8), гадрозаврид (9), дриозаврид (10) и тесцелозаврид (11), орнитопод промежуточного размера. Пситтакозавриды (12) были мелкими цератопсидами, которые, вероятно, ходили на задних ногах. Пахицефалозавриды (13) также ходили на задних ногах и варьировали по размерам от мелких до среднеразмерных.
По мере того, как травоядные из крупных и медлительных превращались в более быстроногих существ меньшего размера, а некоторые вооружились защитными рогами, хищники, которые питались ими, тоже менялись (рис. 14). Первая половина мелового периода была временем взрывного роста разнообразия теропод. Эти жестокие двуногие машины, питающиеся мясом, становились всё крупнее, и, если было возможно, всё более чудовищными. В северном полушарии появились дейнонихозавры, вооружённые огромными серповидными когтями. Представители этой группы включали дромеозавров и троодонов, которые обладали относительно крупными мозгом и глазами, и были снабжены поднимающимися вторыми пальцами задних лап, заканчивающимися изогнутыми когтями, которые могли выбрасываться вперёд, чтобы резать своих жертв. Теризинозавры, также принадлежащие к этой группе, но явно ограниченные Азией*, не просто были большими, но вдобавок каждая их передняя лапа заканчивалась тремя ужасающими, напоминающими косу когтями.
* Остатки теризинозаврид найдены и в Северной Америке, причём ещё до выхода в свет данной книги: в 2001 г. в Нью-Мексико был обнаружен Nothronychus. Но следует помнить, что, несмотря на родство с хищниками, теризинозавры были растительноядными животными. – прим. перев.