
Рис. 16. Условно-рефлекторное воспроизведение ритма сигнального раздражения в ретикулярной формации среднего мозга кошки (отмечено квадратом) (по Н. Иошии и др.):
1 - отметка сигнального звукового раздражителя, 2 - переднее вентральное ядро таламуса, 3 - срединный центр, 4 - ретикулярная формация, 5 - подкрепляющий ритмический световой стимул
На этом основании было высказано предположение, что временные связи условного рефлекса образуются на подкорковом уровне в неспецифических структурах мозгового ствола. Полагали, что при сочетании условного и безусловного раздражителей по коллатералям их специфических путей происходит конвергенция возбуждений в неспецифические структуры ретикулярной формации среднего мозга. Так объясняли образование их связей, а ослабление восходящих активирующих влияний считали причиной ограничения области десинхронизации в коре. Однако наблюдения за динамикой электрической активности корковых и подкорковых структур в процессе выработки и укрепления условного рефлекса показали, что активность неспецифических образований мозгового ствола наиболее выражена в начальных стадиях выработки, когда они вызывают диффузное возбуждение, а по мере упрочения рефлекса на первый план выступают специфические системы на уровне коры (показано в опытах на кошках и собаках).
Деятельность неспецифических структур мозга, в частности восходящей активирующей системы ретикулярной формации среднего мозга, как показали исследования, действительно играет важную роль в образовании временных связей, обеспечивая высокую степень работоспособности высших отделов мозга. Замыкание временных связей является многоуровневым процессом, в который вовлекаются как кора, так и подкорковые образования, причем большую роль в этих процессах играет гиппокамп.
Широкое участие различных структур мозга в образовании временных связей проявляется и в синхронизации ритмов электрической активности корковых областей и многих подкорковых структур. Однако функция большинства из них, особенно неспецифических областей, связана с созданием условий для образования временных связей. Это достигается активацией высших отделов мозга и распространением этого активного состояния на обширные области коры. По мере укрепления связей эти функции отпадают и участие неспецифических областей становится все более ограниченным. Многие электрические показатели этих процессов, резко выраженные в начале выработки рефлекса, в дальнейшем ослабляются и могут исчезнуть. Так происходит с условной реакцией десинхронизации ЭЭГ человека при упрочении условного двигательного рефлекса. Как видно из рис. 17, четкая депрессия альфа-ритма в отведении, соответствующем корковому участку движения руки, по мере увеличения числа сочетаний становится все меньше и при прочном рефлексе совершенно исчезает.
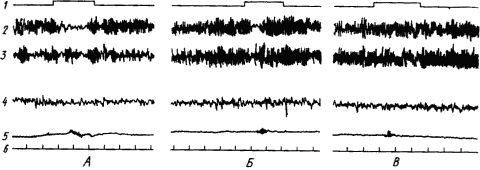
Рис. 17. Изменение ЭЭГ человека при выработке условного двигательного рефлекса на звук метронома. А - после 10 сочетаний; Б - после 28 сочетаний; В - после 80 сочетаний (по А.Б. Когану):
1 - включение метронома, 2, 3 - отведение от левой и правой роландовых борозд, 4 - от височной коры, 5 - электромиограмма общего разгибателя пальцев руки, реагирующей движением, 6 - отметка времени (1 с)
Исследование фоновой ритмики при выработке условных пищевых и оборонительных рефлексов у кроликов в условиях свободного поведения показало учащение колебаний ЭКоГ и укорочение периода условного ответа. При этом наблюдали сходный характер изменений электрической активности при пищевом и оборонительном рефлексах. В специальных опытах с созданием пищевой (голодание) или оборонительной (болевое раздражение) доминанты в коре, ретикулярной формации и лимбических структурах возникали синхронизированные колебания, отражающие высокий уровень неспецифической активации. Эта активация проявлялась не только в учащении импульсов ответов нейрона на условные раздражители, но и в периоды между их предъявлением (рис. 18). Так в деятельности нейронов отражались фазические и тонические компоненты ее перестроек при образовании как пищевых, так и оборонительных условных рефлексов. Однако прямо противоположная реакция в случае пищевого или болевого подкрепления сигнала означает, что проявившаяся в электрической активности неспецифическая активация избирательно действует на структуры мозга, реализующие процессы разного биологического значения. Пищевое подкрепление условного рефлекса придает ему эмоционально-положительный, а оборонительное - эмоционально-отрицательный характер.
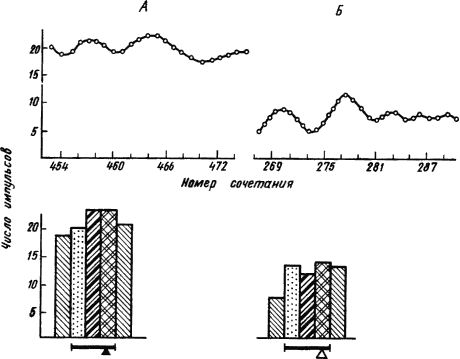
Рис. 18. Тонические и фазические импульсные реакции нейронов гиппокампа при условном пищевом (А) и оборонительном (Б) рефлексах (по Б.И. Котляру, Н.О. Тимофеевой):
А - уровень фоновой активности между сочетаниями, Б - усредненное количество импульсов за 1 с в разные этапы условно-рефлекторной деятельности; чертой отмечено время действия условного раздражителя, треугольником - момент условной реакции
Процессы образования временных связей находят отражение и в сверхмедленных колебаниях потенциалов (СМКП). Так, при выработке у кролика условного оборонительного рефлекса на световой сигнал, сопровождаемый электрокожным подкреплением, наблюдали закономерные изменения СМКП. При этом в начальный период увеличиваются амплитуда и частота СМКП в коре и ряде подкорковых структур, достигая максимума при генерализации рефлекса. Прочное его закрепление сопровождается уменьшением СМКП до полного исчезновения, а угашение рефлекса вновь их вызывает. При выработке условных рефлексов у кроликов происходил сдвиг потенциалов коры вначале в сторону позитивности, а затем негативности, нарастающей по мере укрепления рефлекса. Такую негативизацию рассматривают как показатель деполяризации, обусловливающей местное возбуждение.
Обнаружение связи медленных электрических процессов с психической деятельностью человека (Н.П. Бехтерева, 1966) было исходным для целого направления исследований их динамики, например при обучении иностранным словам или использовании для анализа нейрофизиологических механизмов мнестической деятельности и процесса формирования матрицы долгосрочной памяти. В последнее время СМКП связывают с метаболическими процессами, влияющими на электродный потенциал, а их сдвиги, наблюдаемые при выработке условного рефлекса, обозначают как колебания биоэлектрохимической активности.
Таким образом, при формировании временных связей динамика электрических реакций десинхронизации основной ритмики сверхмедленных потенциалов и усвоения ритма условного раздражителя отражает вначале широкий охват возбуждением неспецифических и специфических систем на корковом и подкорковом уровнях. Он соответствует стадии генерализации условного рефлекса, по мере закрепления которого происходит сосредоточение этих электрических реакций в специфических структурах корковых проекций сигнала и двигательного ответа, а также в подкорковых образованиях.
Вызванные потенциалы. Рассмотренное выше явление усвоения ритмов раздражения представляет собой серии укороченных вызванных потенциалов, в которых полное развитие каждого из них прерывается возникновением следующего. В более полном виде вызванные потенциалы (ВП) с их первичными ответами и поздними компонентами выявляются при относительно редких (1–2/с) раздражениях и могут быть использованы как показатели динамики процессов высшей нервной деятельности. В результате многочисленных исследований изменений ВП в зоне сигнального раздражителя при образовании временных связей накопились разнообразные и во многом противоречивые сведения о закономерностях этих изменений.
Так, в первых исследованиях динамики ВП проекционных зон сигнального раздражителя при выработке условного рефлекса одни авторы отмечали его увеличение, другие - уменьшение, третьи - вообще ставили под сомнение наличие закономерной корреляции между ходом образования условного рефлекса и изменениями ВП. Видимо, причина противоречивых результатов состояла в том, что изменения ВП в чрезвычайной степени зависят от стадии образования условного рефлекса (генерализованного или специализированного, непрочного или упроченного), от локализации отведения (проекция условного или безусловного раздражения, первичные или вторичные сенсорные зоны), от вида, возраста и состояния объекта.
Дальнейшие исследования наглядно показали наличие такой зависимости. Изменения ВП происходят по стадиям формирования условного рефлекса. В большинстве случаев начало образования временной связи может сопровождаться некоторым снижением величины ВП. На стадии генерализации условного рефлекса иррадиация возбуждения в коре мозга проявляется в виде усиления и широкого распространения ВП почти на все области коры и укорочения латентных периодов их ответов на раздражители.