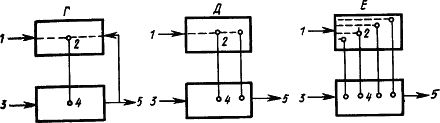
Рис. 11. Различные предположения о структуре временной связи условного рефлекса (объяснение см. в тексте):
1 - условный раздражитель, 2 - корковые структуры, 3 - безусловный раздражитель, 4 - подкорковые структуры, 5 - рефлекторная реакция; прерывистыми линиями показаны временные связи
Рассмотрение процессов замыкания временных связей как центральных звеньев формирования функциональной системы (П.К. Анохин, 1961) относит замыкание к структурам коры, где происходит сопоставление содержания сигнала - афферентный синтез - и результата условно-рефлекторного ответа - акцептор действия (рис. 11, Г).
Изучение двигательных условных рефлексов показало сложную структуру образующихся при этом временных связей (Л.Г. Воронин, 1952). Каждое совершающееся по сигналу движение само становится сигнальным для складывающихся при этом двигательных координаций. Образуются две системы временных связей: на сигнал и на движение (рис. 11, Д).
Наконец, исходя из того, что условные рефлексы сохраняются при хирургическом разделении сенсорных и моторных корковых областей и даже после множественных разрезов коры, а также учитывая, что кора обильно снабжена как входящими, так и выходящими путями, было высказано предположение, что замыкание временных связей может происходить в каждом ее микроучастке между его афферентными и эфферентными элементами, которые активируют центры соответствующих безусловных рефлексов, служащих подкреплением (А.Б. Коган, 1961) (см. рис. 9 и 11, Е). Такое предположение соответствует представлению о возникновении временной связи в пределах анализатора условного раздражителя (О.С. Адрианов, 1953), мнению о возможности "местных" условных рефлексов, замыкающихся внутри проекционных зон (Э.А. Асратян, 1965, 1971), и заключению, что в замыкании временной связи ключевую роль играет всегда афферентное звено (У.Г. Гасанов, 1972).
Нейронная структура временной связи в коре больших полушарий. Современные сведения о микроскопическом строении коры больших полушарий в сочетании с результатами электрофизиологических исследований позволяют с известной степенью вероятности судить о возможном участии тех или иных корковых нейронов в образовании временных связей.
Высокоразвитая кора мозга млекопитающих, как известно, делится на шесть слоев разного клеточного состава. Приходящие сюда нервные волокна заканчиваются большей частью у клеток двух видов. Один из них - это вставочные нейроны, расположенные во II, III и отчасти IV слоях. Их аксоны идут в V и VI слои к крупным пирамидным клеткам ассоциативного и центробежного типа. Это наиболее короткие пути, которые, возможно, и представляют собой врожденные связи корковых рефлексов.
Другой вид клеток, с которыми приходящие волокна образуют наибольшее число контактов, составляют кустовидно ветвящиеся округлые и угловатые короткоотростчатые клетки, часто имеющие звездчатую форму. Они располагаются главным образом в IV слое. Их количество увеличивается с развитием мозга млекопитающих. Это обстоятельство наряду с тем, что звездчатые клетки занимают положение конечной станции для приходящих в кору импульсов, позволяет предположить, что именно звездчатые клетки являются основными воспринимающими корковыми клетками анализаторов и что увеличение их численности в эволюции представляет собой морфологическую основу достижения высокой тонкости и точности отражения окружающего мира.
Система вставочных и звездчатых нейронов может вступать в бесчисленное множество контактов с ассоциативными и проекционными крупными нейронами пирамидной формы, расположенными в V и VI слоях. Ассоциативные нейроны своими аксонами, проходящими через белое вещество, соединяют между собой разные корковые ноля, а проекционные нейроны дают начало путям, соединяющим кору с низшими отделами мозга.
Согласно такому представлению о функциях корковых клеток замыкание временной связи осуществляется в системе звездчатых, вставочных и ассоциативных нейронов. Возможны два типа замыкания. Один из них - это замыкание с участием преимущественно зрительных, слуховых и других экстероцептивных раздражителей, которое происходит с помощью всех указанных нейронов, начиная со звездчатых. Другой - замыкание с участием преимущественно проприо- и интероцептивных раздражителей, происходящее прямо через вставочные нейроны верхних слоев без участия звездчатых нейронов. Предполагают, что именно поэтому некоторые условные рефлексы на проприо- и особенно интероцептивные раздражители, осуществляясь по коротким путям, не вовлекают в деятельность механизмы высших функций мозга человека и, не получая четкого "зеркального" отображения в сознании, проходят помимо него.
Расположение различных корковых нейронов на путях прохождения импульсов и динамика функциональных показателей их активности при образовании условного рефлекса позволяет предположить, что замыкание происходит главным образом на клеточных телах вставочных нейронов. Возможно, особая роль при этом принадлежит нейронам с распространенными рецептивными полями. Существует мнение, что в онтогенезе процессы замыкания зависят от созревания множественных выростов на дендритах нейронов - шипиках.
Известны многочисленные попытки выделить по свойствам нейронов клетки, участвующие в образовании временных связей. Такими свойствами считали высокую частоту фоновой импульсации, большую реакцию на условный раздражитель, способность реагировать на раздражители разной модальности, полисенсорность, высокую лабильность, способность к пластическим перестройкам - избирательную полифункциональность, сходство реакции на условный и безусловный раздражители, ответы преимущественно на подкрепление. На основании опытов главным образом с нервной системой моллюсков возникло представление о ведущей роли в образовании новых форм поведения так называемых командных нейронов, осуществляющих запуск координированных двигательных актов.
Что же происходит с нейронами, участвующими в замыкании временной связи? Давно замечено, что после выработки условного рефлекса возрастает и величина безусловного рефлекса, хотя сила подкрепляющего раздражителя остается прежней. Отсюда следует, что выработка условного рефлекса связана с повышением возбудимости нервных клеток, образующих новый рефлекторный путь. Повышение возбудимости корковых структур, участвующих в образовании условно-рефлекторной связи, можно обнаружить и непосредственно, измеряя пороги их раздражения.
После вживления электродов в область двигательного анализатора (например, кошки), можно измерять порог прямого раздражения и определять изменения возбудимости исследуемых нервных клеток мозга у животного при выработке условного рефлекса. Оказалось, что процесс выработки двигательного условного рефлекса выражается прежде всего стойким повышением возбудимости клеток двигательного анализатора (рис. 12). Только когда возбудимость корковых клеток повышается до известного уровня, появляются двигательные признаки условного рефлекса - сгибание лапы. Каждый очередной сигнал вызывает быстрый дополнительный подъем возбудимости нейронов двигательного анализатора. Если же отменить условный рефлекс, то возбудимость нервных клеток снижается до уровня, предшествующего началу выработки рефлекса. Соответственно изменяются импульсация и ответы на условный раздражитель. При этом меняется деятельность нейронов.
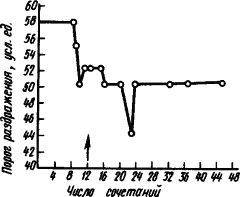
Рис. 12. Изменение порога прямого раздражения сенсомоторной коры при выработке условного рефлекса сгибания лапы у кошки (по Н.И. Николаевой):
стрелкой обозначен момент образования условного рефлекса
Так, полисенсорные нейроны, которые наиболее часто перестраиваются при образовании условного рефлекса, могут стать моносенсорными или не участвовать в условных реакциях. Исходя из того что при разных видах условно-рефлекторной деятельности в разрядах нейронов лобной коры доминируют межимпульсные интервалы каждый раз определенной длительности, было высказано предположение что положительный условный рефлекс и его торможение кодируются различными классами межимпульсных интервалов.